How Scars Form: The Science of Wound Healing
Scar tissue is collagen laid down fast and in parallel — structurally weaker than the basket-weave of normal skin. How the four healing stages work, why keloids and atrophic scars form, and what reduces scarring.
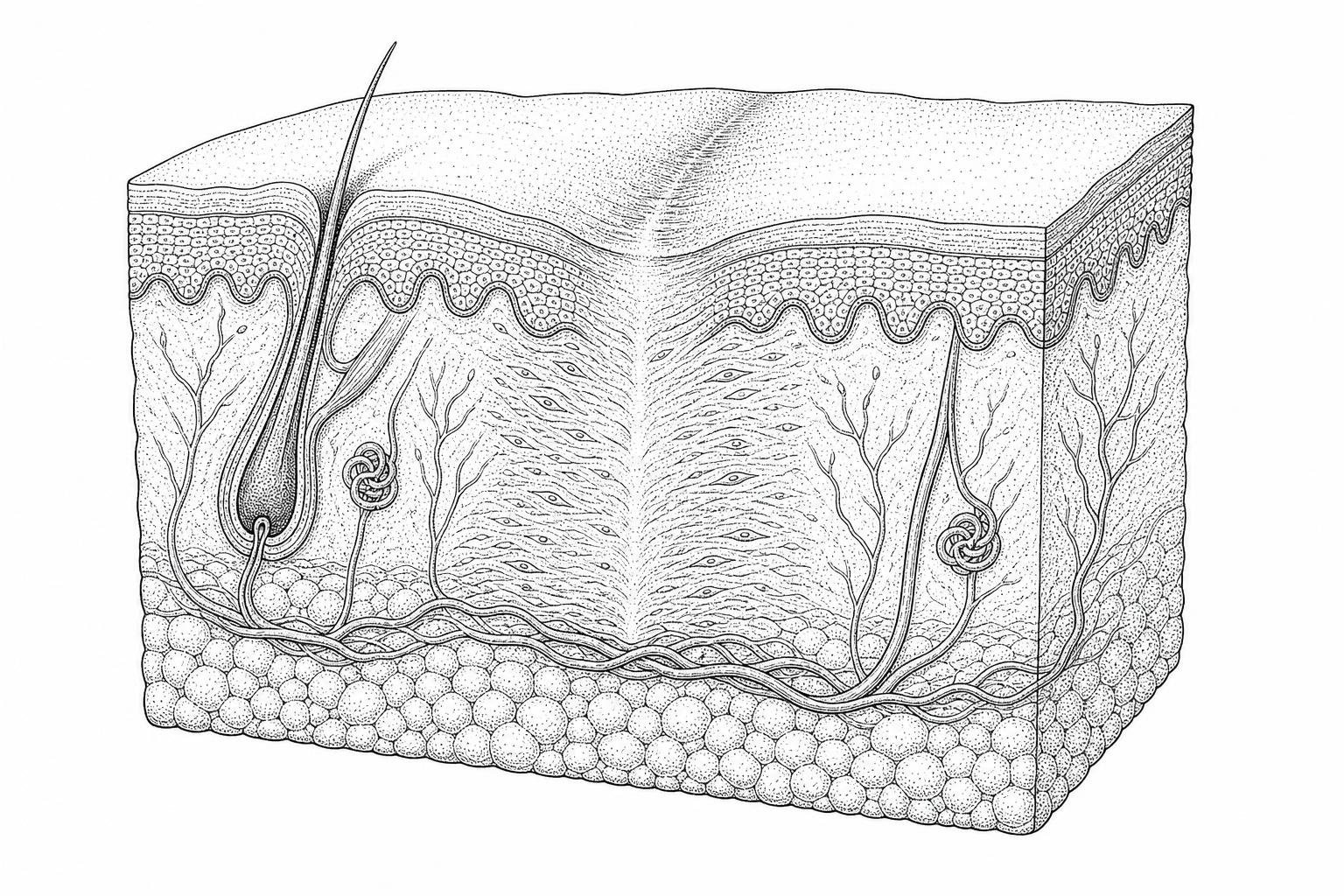
How wounds heal and scars form

How scars form is one of the most common questions people have after an injury, surgery, or a bout of severe acne — and the answer is more complex than most expect.
The short answer:
- Skin is damaged beyond the outer layer (epidermis) into the deeper dermis
- The immune system responds with inflammation to stop bleeding and fight infection
- Fibroblasts migrate to the wound and deposit collagen to close the gap
- New tissue forms — but it is structurally different from the original skin
- Remodeling occurs over months, but the tissue never fully returns to its original architecture
That last point is key. Scar tissue is made of the same protein as normal skin — collagen — but the fibers are arranged differently. Instead of the random, basket-weave pattern of healthy dermis, scar collagen runs in parallel bundles. The result is tissue that is visibly and functionally distinct: firmer, less elastic, and lacking hair follicles, sweat glands, and other skin structures.
Nearly everyone will develop at least one scar in their lifetime. Yet the extent of scarring varies enormously — from a barely visible line to a raised keloid — depending on wound depth, genetics, body location, and how the healing process unfolds at the cellular level.
Understanding the biology behind scar formation is the first step toward managing it more effectively.
This content is for informational purposes only and does not constitute medical advice. Consult a qualified healthcare professional for diagnosis and treatment.
What happens inside a scar
Cutaneous wound healing is a highly coordinated biological response to injury. The primary evolutionary goal of this process is to rapidly restore skin barrier function, preventing exsanguination and pathogenic invasion. However, in adult humans, this rapid sealing mechanism comes at a structural cost: the formation of a fibrotic scar rather than perfect tissue regeneration.
Whether a wound scars depends heavily on the depth of the dermal penetration. Superficial injuries confined strictly to the epidermis heal via simple epithelial cell migration and do not leave a scar. However, once an injury penetrates the basement membrane zone into the papillary and reticular layers of the dermis, the native tissue architecture is disrupted. The body cannot simply regrow the lost dermis in its original configuration. Instead, it initiates a complex reparative cascade that culminates in the deposition of fibrous connective tissue, as outlined in the Scars | Nature Reviews Disease Primers literature.
How scar tissue differs from normal skin
At the microscopic level, healthy skin and scar tissue are fundamentally different. These differences dictate not only how the tissue looks but also how it behaves under physical stress:
- Collagen Alignment: In uninjured skin, collagen fibers (primarily Type I and Type III) are woven in a random, basket-weave pattern that allows the skin to stretch and resist tension in all directions. In scar tissue, the newly deposited collagen fibers are smaller and aligned in tight, parallel bundles running in a single direction.
- Loss of Appendages: Scar tissue lacks specialized skin appendages. Because of the extensive dermal and epidermal damage that occurs during injury, sweat glands, sebaceous glands, and hair follicles do not regenerate within within mature scar tissue.
- Absence of Rete Pegs: Healthy skin features downward epithelial projections called rete pegs that interlock with the dermal papillae below, preventing the layers from shearing. Scars lack these structures, making them far more susceptible to mechanical shear forces.
- Elastin Deficiency: Elastic fibers, which give healthy skin its springiness, are severely deficient or entirely disorganized in scar tissue. Research indicates that elastin is rarely detected in scars younger than three months and remains permanently deficient in deep human wounds.
- Tensile Strength Limits: Normal skin has a high tensile strength comparable to steel on a per-weight basis. A healing surgical wound or deep scar takes about six weeks to reach 50% of original strength, and eight to ten weeks to reach approximately 80%. Even after a year of remodeling, mature scar tissue never exceeds 80% to 90% of the original tensile strength of uninjured skin.
How Scars Form During the Four Stages of Wound Healing
The transition from a fresh wound to a mature scar occurs across four highly overlapping, temporally regulated phases: hemostasis, inflammation, proliferation, and remodeling. A detailed review of these phases can be found in Healing Mechanisms in Cutaneous Wounds: Tipping the Balance.
1. Hemostasis (Minutes to Hours)
Immediately following injury, exposed subendothelial collagen activates blood platelets. These platelets aggregate to form a primary hemostatic plug and release clotting factors that initiate the coagulation cascade, resulting in a stable fibrin clot. Platelets also degranulate, releasing vital growth factors—such as Transforming Growth Factor-beta (TGF-β), Platelet-Derived Growth Factor (PDGF), and Epidermal Growth Factor (EGF)—which act as chemical beacons to recruit inflammatory cells.
2. Inflammation (Days 1 to 3, lasting up to 2 weeks)
Within 24 hours, neutrophils migrate to the wound site to clear cellular debris and bacteria via phagocytosis. They are typically depleted within two to three days via apoptosis. Monocytes then enter the tissue and mature into macrophages. Macrophages act as the master orchestrators of healing; they secrete cytokines, coordinate angiogenesis (the growth of new blood vessels), and transition the wound microenvironment from a pro-inflammatory state to a pro-healing, proliferative state.
3. Proliferation (Days 4 to 21)
This phase is characterized by the formation of highly vascularized granulation tissue. Fibroblasts migrate into the wound space, driven by local chemotactic signals. Once there, they rapidly synthesize extracellular matrix (ECM) proteins, primarily collagen Type III, fibronectin, and hyaluronic acid. The fundamental role of amino acids and structural proteins during this phase is discussed in the clinical review on Protein Synthesis Wound Healing. Concurrently, endothelial cells form new capillary networks to supply oxygen, while keratinocytes migrate across the granulation tissue to re-establish the epithelial barrier.
4. Remodeling (Three Weeks to Two Years)
Also known as the maturation phase, this is where the definitive scar is shaped. The highly cellular, vascular granulation tissue is gradually replaced by an acellular, avascular collagenous matrix. Under the influence of matrix metalloproteinases (MMPs) and their tissue inhibitors (TIMPs), the initially deposited Type III collagen is progressively degraded and replaced by stronger Type I collagen. The collagen bundles are slowly cross-linked and realigned along local mechanical stress lines.
What drives excess scar tissue
When the wound healing cascade is altered, normal tissue repair can shift toward pathological fibrosis. This transition is governed by a delicate balance of cellular signaling pathways, mechanical cues, and specific cell lineages, as detailed in Chapter 3 Scar Formation: Cellular Mechanisms.
The cells that build scar tissue
Historically, it was assumed that all dermal fibroblasts at a wound site behaved identically and arose de novo from local connective tissue. Modern single-cell lineage tracing has disproven this, revealing a high degree of fibroblast heterogeneity.
Crucially, research has identified that all fibrotic scars in adult mammals derive from a specific fibroblast lineage expressing the Engrailed-1 gene (ENF lineages). In contrast, Engrailed-1 negative fibroblasts (ENFs) are associated with non-scarring, regenerative healing.
Furthermore, groundbreaking research has challenged the traditional paradigm of wound repair by demonstrating that scars do not form entirely de novo at the surface of the wound. Instead, a significant portion of scar tissue originates from the fascia—the deep, connective tissue layer beneath the skin.
The fascia contains specialized, prefabricated kits of "sentry" fibroblasts embedded in a highly mobile, sealant-like matrix jelly. Upon injury, these fascia fibroblasts drag this prefabricated matrix jelly upward into the open wound gap to rapidly plug the defect. Experimental models show that when these fascia-derived fibroblasts are selectively removed or inhibited, the matrix fails to incorporate into the wound, and only abnormal, poorly healed wounds result. This discovery has profound implications for the study of Scar Science, shifting focus toward therapeutic targeting of the deep fascial layers.
How wounds tighten as they heal
The primary cellular driver of both physiological wound closure and pathological scarring is the myofibroblast. Myofibroblasts are specialized, highly contractile cells that share characteristics with both fibroblasts and smooth muscle cells.
The differentiation of a standard fibroblast into a fully active myofibroblast occurs in a stepwise fashion:
- Proto-Myofibroblast Formation: Under the influence of mechanical tension and the deposition of ED-A fibronectin, local fibroblasts develop cytoplasmic stress fibers but do not yet express contractile proteins.
- Full Differentiation: The synergy of high mechanical stress and local activation of Transforming Growth Factor-beta 1 (TGF-β1) drives the proto-myofibroblast to express alpha-smooth muscle actin (α-SMA) within its stress fibers.
Once fully differentiated, myofibroblasts physically bind to the surrounding extracellular matrix via specialized integrin receptors and connect to one another through adherens junctions. This allows them to act like a coordinated cellular network (a syncytium), pulling the edges of the wound together through sustained contraction. In a rat model, myofibroblasts can constitute up to 70% of the active fibroblasts present in the forming scar tissue.
In normal, physiological healing, once the wound is successfully closed and the mechanical tension is relieved, these myofibroblasts undergo programmed cell death (apoptosis). However, if the inflammatory phase is prolonged or if mechanical tension remains high, these myofibroblasts persist. They continue to synthesize excessive Type I collagen and fibronectin, leading to the dense, contracted, and raised tissue characteristic of pathological scarring.
The main types of scars
The clinical presentation of a scar is determined by the balance between extracellular matrix synthesis and degradation during the remodeling phase. When this balance is disrupted, it leads to distinct fibroproliferative disorders or tissue-deficit scars, as explored in Scarring Skin: Mechanisms and Therapies.
Hypertrophic Scars vs. Keloids
Hypertrophic scars and keloids represent two distinct forms of excessive dermal fibrosis. While both are characterized by overactive fibroblasts and excessive collagen deposition, they differ significantly in their clinical behavior, histology, and response to treatment.
The biological stages of these raised lesions are detailed further in the Keloid Scar Formation Stages guide.
| Clinical & Biological Feature | Hypertrophic Scars | Keloid Scars |
|---|---|---|
| Growth Boundaries | Remain strictly within the margins of the original wound. | Extend beyond the original wound margins, invading healthy surrounding skin. |
| Onset Timeline | Typically appear rapidly (within 4 to 8 weeks) post-injury. | Can appear months to years after minor or unnoticed trauma. |
| Anatomical Site Preference | Commonly form over high-tension areas (joints, shoulders, knees). | Favor the earlobes, chest, shoulders, and upper back. |
| Histological Structure | Contain organized, parallel bundles of Type III collagen. | Contain thick, disorganized, glassy "keloidal" Type I and III collagen. |
| Genetic & Ethnic Risk | Show no strong racial predilection; related to wound tension. | Highly prevalent in dark-skinned populations and red-haired individuals. |
| Resolution Behavior | Often regress spontaneously or flatten over several years. | Rarely regress spontaneously; high recurrence rates (up to 45%) after excision. |
Atrophic scars and stretch marks
In contrast to hypertrophic scars, atrophic scars are characterized by a net loss of dermal collagen and subcutaneous fat. These scars typically form when an intense, localized inflammatory response—such as that seen in cystic acne, varicella (chickenpox), or deep insect bites—destroys the underlying dermal matrix before sufficient collagen synthesis can occur.
Atrophic scars are generally categorized into three clinical shapes:
- Icepick Scars: Deep, narrow (under 2mm), vertical tracts that extend into the deep dermis or subcutaneous tissue.
- Boxcar Scars: Round to oval depressions with sharply defined, vertical margins.
- Rolling Scars: Wide, shallow depressions caused by abnormal fibrous anchoring of the dermis to the subcutaneous tissue, creating a wave-like appearance on the skin surface.
Striae Distensae (Stretch Marks) represent a unique form of dermal scarring. They occur when rapid stretching of the skin—such as during pregnancy, rapid weight gain, or adolescent growth spurts—exceeds the elastic limits of the dermis. This mechanical stress causes structural failure of the dermal extracellular matrix, leading to the rupture of elastic fibers and the localized degradation of collagen. The resulting scars appear as linear, thin, atrophic bands that lack normal skin appendages and elasticity.
What else affects how scars heal
Wound healing does not occur in a cellular vacuum. It is heavily influenced by physical forces, nervous system signaling, and the unique biochemical environments of different life stages.
How Mechanical Tension Shapes Scar Formation
Cells are highly sensitive to their physical environment. The field of mechanobiology has demonstrated that physical forces are translated into biochemical signals through a process called mechanotransduction. The clinical implications of these forces are detailed in the Mechanobiology of Cutaneous Scarring - Textbook on Scar Management - NCBI Bookshelf.
When a healing wound is subjected to mechanical stretching, specific cell-surface receptors (such as integrins and stretch-activated ion channels) are stimulated. This mechanical loading triggers several cellular responses:
- It activates latent TGF-β1 stored within the extracellular matrix, further driving myofibroblast differentiation.
- It upregulates the expression of pro-fibrotic genes, leading to increased collagen synthesis.
- It suppresses myofibroblast apoptosis, causing these contractile cells to persist and continue depositing matrix tissue.
This explains why hypertrophic scars frequently form near highly mobile joints (such as the shoulder, elbow, and knee) or across the chest, where respiration constantly stretches the skin. Conversely, areas where the skin is stabilized by underlying bone or experiences minimal movement, such as the scalp or anterior lower leg, rarely develop thick, pathological scars.
Furthermore, the physical shape of a keloid is often dictated by the direction of these physical forces. For example, chest keloids frequently expand horizontally into a "butterfly" or "crab's claw" shape, directly aligning with the lateral tension lines created by daily arm movements.
How nerves affect scar healing
The cutaneous nervous system plays an active role in modulating wound repair. The skin is richly innervated by sensory nerve fibers, specifically nociceptors (pain-sensing fibers).
During injury, these sensory fibers release potent neuropeptides—such as Substance P (SP) and Calcitonin Gene-Related Peptide (CGRP)—directly into the wound microenvironment. This pathway is explored in depth in a 2024 review of neuropeptides in cutaneous wound healing.
These neuropeptides trigger a cascade known as neurogenic inflammation:
- They act directly on local vascular endothelial cells to increase permeability, facilitating the recruitment of inflammatory cells.
- They stimulate mast cells to degranulate, releasing histamine and other pro-inflammatory mediators.
- They interact with local dermal fibroblasts, promoting their proliferation, migration, and synthesis of collagen.
Clinical observations support this neuro-inflammatory link. Pathological scars, particularly keloids, show a significantly higher density of sensory nerve fibers compared to normal skin. This hyper-innervation correlates directly with the chronic itching, hypersensitivity, and pain frequently reported by patients. Conversely, denervated skin (such as in areas of peripheral neuropathy or surgical nerve damage) often displays altered wound healing kinetics and reduced scarring, highlighting the direct role of active nerve signaling in driving the fibrotic response.
Why babies in the womb heal without scars
One of the most compelling areas of regenerative medicine is the study of mammalian fetal wound healing. Up to approximately 24 weeks of human gestation, cutaneous wounds heal through true regeneration, restoring normal skin architecture—including hair follicles, sweat glands, and a perfect basket-weave collagen matrix—without any trace of a scar.
This scarless regenerative capacity is due to several key biological differences between fetal and adult tissues:
- Attenuated Inflammatory Response: Fetal wounds exhibit a minimal inflammatory response. The levels of pro-inflammatory cytokines (such as IL-6 and IL-8) are low, while the levels of the potent anti-inflammatory cytokine Interleukin-10 (IL-10) are high. IL-10 downregulates collagen Type I expression and limits the recruitment of inflammatory cells.
- Extracellular Matrix Composition: The fetal extracellular matrix is exceptionally rich in hyaluronic acid (HA). This high-HA environment promotes rapid fibroblast migration and prevents the dense, rigid collagen cross-linking seen in adult scars.
- Fibroblast Heterogeneity: Fetal skin contains a high proportion of Engrailed-1 negative fibroblasts (ENFs) and CD26- fibroblasts. These cells deposit collagen in a highly organized, random basket-weave pattern. Fetal fibroblasts also resist differentiating into contractile, α-SMA-positive myofibroblasts, avoiding the tissue contraction that characterizes adult scarring.
- Absence of Hemostasis: Because the fetus exists in a sterile, fluid-filled amniotic environment, fetal wounds do not require a rapid, fibrin-heavy hemostasis phase. Re-epithelialization begins almost immediately without the formation of a dense fibrin clot.
How to prevent and treat scars
While complete scarless healing remains a major goal of regenerative medicine, several evidence-based clinical interventions are currently utilized to prevent and manage abnormal scarring.
Treatments that work today
Modern scar management relies on therapies designed to normalize the wound microenvironment, control mechanical tension, and downregulate fibroblast activity. A comprehensive overview of these modalities can be found in the Scar Revision Treatment Complete Guide.
- Topical Silicone Therapy: Silicone gel sheeting and topical silicone gels represent the gold standard for non-invasive scar prevention and treatment. While the exact molecular mechanism remains under study, clinical evidence suggests that silicone sheets work primarily by restoring the stratum corneum barrier. This reduces transepidermal water loss (TEWL), signaling to the underlying fibroblasts to downregulate collagen production. Additionally, the physical sheet provides slight mechanical stabilization, reducing lateral skin tension.
- Corticosteroid Injections: Intralesional injections of corticosteroids (primarily triamcinolone acetonide) are highly effective for raised hypertrophic scars and keloids. Corticosteroids act by suppressing the local inflammatory response, inhibiting fibroblast proliferation, and reducing the synthesis of collagen and glycosaminoglycans. They also upregulate collagenase (MMPs), which helps break down existing excess collagen.
- Pressure Garment Therapy: Often utilized in burn reconstruction, pressure garments apply continuous mechanical compression (typically 15 to 25 mmHg) to the healing tissue. This physical compression induces local hypoxia, which reduces cellular metabolism, limits capillary growth, and accelerates the apoptosis of overactive myofibroblasts.
- Surgical Scar Revision: For mature, contractured, or highly visible scars, surgical intervention may be required. Techniques such as Z-plasty or W-plasty are designed to physically redirect the scar's direction, aligning it with natural skin tension lines (Langer's lines) to minimize post-operative mechanical stress.
The future of scarless healing
Recent advances in molecular biology have opened new pathways for preventing scars at the cellular level. Several promising therapies are currently undergoing preclinical and clinical evaluation:
- Mechanical Stress Shielding: Utilizing specialized adhesive dressings designed to absorb lateral skin tension around a fresh incision, this approach prevents mechanical forces from activating local mechanosensors. By shielding the wound from stretching, it prevents the activation of pro-fibrotic pathways.
- Verteporfin (Engrailed-1 Inhibitor): Verteporfin, a small molecule FDA-approved for macular degeneration, has recently been shown to block the mechanotransduction pathways that drive fibroblasts to express the Engrailed-1 gene. Animal studies have shown that applying Verteporfin to fresh wounds can prevent fibrotic scar formation, resulting in regenerated skin complete with functional hair follicles and sweat glands.
- CD26 Inhibition: CD26 is a cell-surface marker associated with the lineage of fibroblasts responsible for scar deposition. Inhibiting CD26 activity (using compounds like diprotin A) has shown promise in reducing fibrotic collagen deposition and promoting a more organized, regenerative tissue repair process.
To evaluate individual healing risk and track scar maturation, clinicians utilize standardized tools. These methods are detailed in the Scar Assessment guide.
Common Questions About How Scars Form
Why do scars lack hair follicles and sweat glands?
Hair follicles and sweat glands are complex epidermal appendages that develop during embryonic morphogenesis. This development requires a highly coordinated exchange of signals between the epidermis and the deep dermis.
When a deep dermal injury occurs, this delicate signaling network is permanently disrupted. The rapid, reparative deposition of parallel collagen bundles focuses on sealing the skin barrier quickly rather than slowly rebuilding these complex structures. As a result, mature scar tissue lacks the specialized stem cell niches required to regenerate hair follicles and sweat glands.
Can a scar completely disappear over time?
A true scar that has penetrated the dermis will never completely disappear. While the remodeling phase can continue for up to two years—during which the scar may fade from red to pale, soften, and flatten—the underlying parallel collagen structure is permanent. The tissue will never fully regain the random basket-weave architecture of uninjured skin, and its tensile strength will permanently remain at roughly 80% to 90% of its original capacity.
Why are some skin types more prone to keloids?
The predisposition to keloid scarring has a strong genetic component. Keloids are significantly more common in individuals with darker skin tones (including those of African, Asian, and Hispanic heritage) and are also observed with higher frequency in red-haired individuals.
Research indicates that these populations often exhibit genetic variations that lead to hyper-reactive fibroblasts, increased sensitivity to TGF-β, and a prolonged, low-grade inflammatory response to minor skin trauma. In many cases, this predisposition is inherited across generations within families.
Key Takeaways and the Future of Scar Treatment
The biological process of how scars form is a complex, multi-phased survival mechanism. While the rapid deposition of parallel collagen fibers successfully restores the skin barrier, it leaves a permanent structural mark.
From the deep fascial origin of scar-forming fibroblasts to the profound influence of mechanical tension and neurogenic inflammation, modern science is steadily uncovering the precise molecular cues that dictate whether a wound heals via fibrotic repair or regenerative scarless healing.
Understanding these pathways allows for more effective prevention, better clinical management, and targeted therapeutic interventions. To evaluate your healing progress or assess a specific scar, consider utilizing the clinical Scar Assessment tool.
Works Cited
- Singer, A.J. "Healing Mechanisms in Cutaneous Wounds: Tipping the Balance." Tissue Engineering Part B: Reviews, 2022.
- Darby, I.A., and A. Desmoulière. "Scar Formation: Cellular Mechanisms." Textbook on Scar Management, 2020.
- Lin, X., and Y. Lai. "Scarring Skin: Mechanisms and Therapies." International Journal of Molecular Sciences, 2024.
- Ogawa, R. "Mechanobiology of Cutaneous Scarring." Textbook on Scar Management, 2020.
- Xing, L., et al. "The Role of Neuropeptides in Cutaneous Wound Healing: A Focus on Mechanisms and Neuropeptide-Derived Treatments." Frontiers in Bioengineering and Biotechnology, 2024.
- Jeschke, M.G., et al. "Scars." Nature Reviews Disease Primers, 2023.
This content is for informational purposes only and does not constitute medical advice. Consult a qualified healthcare professional for diagnosis and treatment.